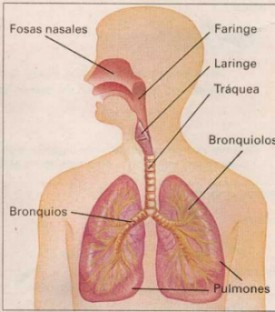
La respiración es el proceso por el cual ingresamos aire (que contiene
oxígeno) a nuestro organismo y sacamos de él aire rico en dióxido de carbono.
Un ser vivo puede estar varias horas sin comer, dormir o tomar agua, pero
no puede dejar de respirar más de tres minutos. Esto grafica la importancia
de la respiración para nuestra vida.
El sistema respiratorio de los seres humanos está formado por: Las vías respiratorias: son las fosas nasales, la faringe,
la laringe, la tráquea, los bronquios y los bronquíolos. La boca también es,
un órgano por donde entra y sale el aire durante la respiración.
Las fosas nasales son dos cavidades situadas encima
de la boca. Se abren al exterior por los orificios de la nariz (donde reside el sentido del olfato) y se comunican
con la faringe por la parte posterior. En el interior de las fosas nasales
se encuentra la membrana pituitaria, que calienta y humedece el aire que inspiramos.
De este modo, se evita que el aire reseque la garganta, o que llegue muy frío
hasta los pulmones, lo que podría producir enfermedades. No confundir esta membrana pituitaria con la glándula pituitaria o hipófisis.
(Ver: La nariz y el olfato)
La faringe se
encuentra a continuación de las fosas nasales y de la boca. Forma parte
también del sistema digestivo. A través de ella pasan el alimento que
ingerimos y el aire que respiramos.
La laringe está
situada en el comienzo de la tráquea. Es una cavidad formada por cartílagos
que presenta una saliente llamada comúnmente nuez. En la laringe
se encuentran las cuerdas vocales que, al vibrar, producen la voz.
La tráquea es
un conducto de unos doce centímetros de longitud. Está situada delante
del esófago.
Los bronquios son los dos tubos en que se divide la
tráquea. Penetran en los pulmones, donde se ramifican una multitud de veces,
hasta llegar a formar los bronquiolos.
La digestión es el proceso de transformación de los alimentos,
previamente ingeridos, en sustancias más sencillas para ser absorbidos.
La digestión ocurre tanto en los organismos pluricelulares como en las células, (ver digestión intracelular). En este proceso participan diferentes tipos de enzimas.
El sistema o aparata digestivo , es muy importante en la digestión ya que los organismos heterotrofos dependen de fuentes externas de materias primas y energía para crecimiento, mantenimiento y funcionamiento. El alimento se emplea para generar y reparar tejidos y obtención de energía. Los organismos autrótofos (las plantas, organismos fotosinteticos), por el contrario, captan la energía lumínica y la transforman en energía química, utilizable por los animales.
En cada paso de la conversión energética de un nivel a otro hay una
pérdida de materia y energía utilizable asociada a la mantención de
tejidos y también a la degradación del alimento en partículas más
pequeñas, que después se recontituiran en moléculas tisulares más complejas.
También es el proceso en que los alimentos al pasar por el sistema digestivo son transformados en nutrientes y minerales que necesita nuestro cuerpo.
La digestión es el conjunto de mecanismos por los
cuales se produce la degradación de los alimentos y su transformación en
moléculas de tamaño reducido. El proceso supone una rotura enzimática
tanto de glúcidos, como de proteínas y lípidos, por medio de enzimas de la
saliva, jugo gástrico, jugo pancreático y células de las vellosidades
intestinales. Los lípidos requieren además una solubilización micelar por
medio de los ácidos biliares de la bilis.
Los productos finales de la digestión deben a
continuación atravesar la pared del tubo digestivo y pasar a la sangre
(absorción), para ser posteriormente distribuídos a todas las células del
organismo. La absorción intestinal se caracteriza por ser un proceso
rápido, de tal forma que la digestión y absorción de una comida compleja pueden
completarse en menos de tres horas. Es, además, un proceso
extraordinariamente eficaz; por ejemplo, la absorción de lípidos y proteínas es
superior al 95 %.
Aunque en el estómago puede absorberse
alcohol y
algún otro tipo de moléculas (ciertos fármacos del tipo de la aspirina),
sólo
lo hacen en cantidades muy reducidas. Los procesos de absorción tienen
lugar fundamentalmente en el intestino delgado y en el intestino grueso.
El principal lugar de absorción es el intestino delgado, aunque el agua
y las
sales pueden hacerlo también en el intestino grueso.
6.1 Superficie de absorción
Intestino
delgado
Superficie (m2)
Incremento relativo
Tubo liso
0,33
1
Pliegues de Kerckring
1
x 3
Vellosidades
10
x 30
Microvellosidades (borde en cepillo)
300
x 1000
La transferencia de sustancias a través de la
pared del intestino es posible gracias a una estructura especialmente
adaptada. La superficie interna es extraordinariamente grande
(superior a los 300 m2), lo que facilita en
gran medida el paso de sustancias a través de la misma y, por tanto, el proceso
de absorción. Esto es posible, en primer lugar, gracias a la existencia
de una serie de pliegues que incrementan tres veces la superficie (pliegues
de Kerckring); y, en segundo lugar, a la presencia de un gran número de
pequeñas vellosidades que la aumentan aún más.
Por último, las
células de la mucosa intestinal tienen un borde en cepillo, constituido
por un gran número de microvellosidades que hacen todavía mayor la
superficie de absorción. Estas células denominadas enterocitos,
se forman a partir de células precursoras en las profundidades de las
criptas
del Lieberkühn y luego van emigrando hacia el extremo de la vellosidad.
Los enterocitos tienen una vida media de unos pocos días y, cuando
envejecen,
se van descamando a la luz intestinal. El epitelio intestinal se renueva
íntegramente en 6 días.
6.2 Digestión y absorción de glúcidos
El consumo diario de glúcidos en los países
occidentales es de unos 250‑800 g. Más del 50% se encuentran en forma de almidón, y en menores proporciones
como los disacáridos sacarosa y lactosa y los monosacáridos glucosa y fructosa.
La digestión de los hidratos de carbono se inicia
por medio de la ptialina de la saliva, y continúa en el intestino
delgado por la acción de la amilasa pancreática. Ambas actúan
sobre los enlaces (α 1- 4) internos de las moléculas de almidón, dando lugar a
la formación de maltosa (dímeros de glucosa), maltotriosa (trímeros de glucosa)
y dextrinas (que contienen los puntos de ramificación del almidón). Dado
que los glúcidos sólo pueden absorberse en forma de monosacáridos, los
productos de la digestión o los disacáridos de la dieta tienen que continuar
siendo desintegrados en la membrana del borde en cepillo, que contiene
diversos tipos de oligosacaridasas (maltasas, lactasas, sacarasas e
isomaltasas o dextrinasas). La acción de estas enzimas va a dar lugar a
la formación de los monosacáridos glucosa, galactosa y fructosa.
La absorción de los monosacáridos glucosa y
galactosa, tiene lugar por un mecanismo común dependiente de sodio (transporte
activo secundario), dándose fenómenos de inhibición competitiva entre
ambas. Una vez que ambos monosacáridos se encuentran concentrados en el
interior celular, pueden ya penetrar, por difusión facilitada, a través
de la membrana basolateral, y dirigirse a la sangre. A diferencia de la
glucosa y la galactosa, la absorción de la fructosa se lleva a cabo mediante un
mecanismo de difusión pasiva y, una vez dentro de la célula, se produce
una transformación parcial en glucosa antes del paso a la sangre.
6.3 Digestión y absorción de proteínas
La ingestión diaria de proteínas es de
aproximadamente 70‑90 g, siendo sus fuentes
principales la carne y los productos lácteos. Además de la fuente dietética,
una parte importante de las proteínas que llegan al intestino proceden también
de las secreciones digestivas (25%) o la descamación de células epiteliales
(25%).
La digestión de las proteínas se inicia por
acción de la pepsina del estómago, dando lugar a la formación de
polipéptidos, oligopéptidos y algunos aminoácidos. La digestión se
continúa en el intestino gracias a las proteasas del jugo pancreático (tripsina,
quimotripsina, carboxipeptidasas, colagenasa y elastasa), formándose
ya oligopéptidos (30%) y diferentes aminoácidos (70%).
Al igual que ocurría con los glúcidos, la
digestión se completa a nivel del borde en cepillo de las células intestinales
o enterocitos. Estos con tiene una serie de aminopeptidasas
orientadas hacia el exterior de la membrana, que hidrolizan la mayor parte de
los péptidos, liberando aminoácidos. Una pequeña parte de los péptidos
pueden, no obstante, pasar al interior de las células a través de un
transportador ligado a H+ y se hidrolizan a
aminoácidos por medio de peptidasas citoplasmáticas. Los mecanismos de
absorción de los aminoácidos son transporte activo secundario acoplado al
sodio, existiendo cuatro tipos de transportadores distintos, para los
aminoácidos neutros, básicos, ácidos y uno específico para prolina e
hidroxiprolina.
La mayor parte de los productos de la digestión
de las proteínas se absorben en el intestino delgado. Al intestino grueso
sólo llegan pequeñas cantidades que serán catabolizadas por la flora
intestinal. Es importante señalar que, aunque en proporciones
muy pequeñas, también es posible la absorción intestinal de proteínas por
mecanismos de pinocitosis. La importancia nutritiva es mínima, pero sí
puede tener interés al desencadenar una respuesta inmunológica.
6.4 Digestión y absorción de lípidos
El consumo diario de lípidos es de unos 60‑100
g. En su mayor parte son triglicéridos y sólo una pequeña porción se encuentra
en forma de lecitinas, ésteres de colesterol o vitaminas liposolubles.
6.4.1 Emulsificación, digestión e
incorporación a las micelas
La solubilización sólo es posible por
incorporación a las micelas de la bilis. Cuando la bilis se mezcla con
las gotitas de lípidos en el intestino, los lípidos se absorben en las micelas
y así se mantienen estables pasando de formar parte de gotas cuyo diámetro era
de 0,5 a 1 μ, a micelas cuyo diámetro es de 4 a 6 nm (aproximadamente 1.000
veces más pequeñas).
La digestión de los lípidos se lleva a cabo a
nivel de intestino delgado gracias a la presencia de las enzimas lipolíticas
del páncreas. La lipasa pancreática, es la más importante, desdobla los triglicéridos en monogliceridos y ácidos grasos; también parece
existir una lipasa gástrica, capaz de digerir triglicéridos de cadena corta,
pero su actividad es muy reducida. La fosfolipasa disocia las lecitinas
en lisolecitinas y ácidos grasos. La colesterol‑ésterhidrolasa
hidroliza el colesterol esterificado, originando ácidos grasos y colesterol
libre.
Al mismo tiempo, la lipasa se absorbe también,
manteniéndose anclada a los ácidos biliares gracias a una proteína, la colipasa
pancreática. Entonces se produce la hidrólisis de los triglicéridos, con
formación de monoglicéridos y ácidos grasos, que se incorporan a las micelas ya
que los productos de la hidrólisis de los lípidos son compuestos insolubles en
el medio acuoso intestinal.
6.4.2 Entrada al enterocito o célula
epitelial intestinal
Una vez producida la incorporación a las micelas
mixtas, los productos de la digestión de los lípidos pueden ya ponerse en
contacto con las microvellosidades y absorberse a través de la membrana celular
por difusión. Para penetrar en el interior de los enterocitos, las
moléculas lipídicas difunden primero a la zona de líquido que rodea a éstos y
luego penetran a través de la membrana epitelial. Las micelas difunden
entonces en sentido retrógrado y vuelven a absorber nuevos lípidos, que son
transportados hacia las células de las vellosidades.
La absorción intestinal de los lípidos es un
proceso muy eficaz. Más del 95% de los mismos se recuperan, fundamentalmente a
nivel duodenal, y sólo una pequeña cantidad se pierde cada día a través de las
heces.
6.4.3 Metabolismo celular y formación de
quilomicrones
Una vez en el interior de las células
intestinales, los productos de la digestión de los lípidos se unen a una
proteína transportadora de bajo peso molecular, la cual los lleva hasta el
retículo endoplasmático liso. En éste tiene lugar la resíntesis de
triglicéridos, la de lecitinas y la de colesterol esterificado.
Los diferentes lípidos se agrupan posteriormente
y se rodean de una cubierta de betalipoproteínas formadas en el aparato de
Golgi, dando lugar a la aparición de los quilomicrones. Su composición
aproximada sería: 87% de triglicéridos, 9% de fosfolípidos y colesterol libre,
3% colesterol esterificado y 1% de vitaminas liposolubles y proteínas.
6.4.4 Recogida del sistema linfático
Estos quilomicrones abandonan la célula, por
exocitosis, a través de la membrana lateral y se dirigen a los quilíferos (o
lactóforos) centrales de las vellosidades y al interior del sistema
linfático. Los quilomicrones presentan un tamaño lo bastante grande (100
nm) como para dar a la linfa, e incluso al plasma circulante, un aspecto
lechoso después de una comida rica en lípidos. Una excepción al mecanismo
descrito son los ácidos grasos de cadena corta y media, que abandonan las
células sin esterificación y sin incorporarse a los quilomicrones, pasando por
simple difusión a los capilares sanguíneos.
Los triglicéridos de cadena media (TGCM), a
diferencia de los de cadena larga y corta, penetran en la célula de forma
directa, sin disociación previa por medio de la lipasa. Un 30% de los
TGCM pueden ser absorbidos sin hidrólisis previa; los que son hidrolizados lo
hacen por las lipasas pancreáticas, formando rápidamente micelas. Una vez
en el interior de la célula no se reesterifican sino que pasan en seguida a la
circulación venosa portal.
6.4.5 Transporte de los lípidos en sangre
Tipo
Origen
Destino
Lípidos principales
Función
Quilomicrón
Intestino
Células
TG y otros
Transporte de lípidos de la dieta.
VLDL
Hígado
Células
TG y colesterol
Transporte de lípidos endógenos.
LDL
Vasos (resto de VLDL)
Hígado
Colesterol
Transporte colesterol.
HDL
Hígado e intestino
Hígado y células con alto uso de colesterol
Colesterol
Elimina y degrada el colesterol.
A través de la linfa se incorporan a sangre. En
las células endoteliales, por medio de la lipoproteín-lipasa se hidrolizan los
triglicéridos de los quilomicrones, dando ácidos grasos libres y glicerol que
serán usados por las células. Los restos del quilomicrón con elevado contenido
en colesterol son llevados al hígado donde son absorbidos por endocitosis. El
colesterol y triglicéridos sintetizados en el hepatocito son secretados a
sangre unidos a apoproteínas formando complejos denominados VLDL (lipoproteínas
de muy baja densidad), que permiten el transporte de triglicéridos a diferentes
órganos. Al perder triglicéridos las VLDL pasan a LDL y el excedente de
colesterol se reenvía al hígado unido a proteínas formando HDL.
6.5 Absorción de agua e iones
El agua que pasa por el intestino delgado,
aproximadamente unos 9 litros diarios (2 litros procedentes de la alimentación
y unos 7 litros procedentes de las secreciones digestivas), se reabsorbe en más
de un 80% en el intestino delgado. El resto lo hace en el intestino
grueso, de tal modo que sólo una pequeña cantidad, alrededor del 1%, se
elimina con las heces. Los desplazamientos del agua tienen lugar por mecanismos
pasivos osmóticos, y se relacionan normalmente con el transporte de solutos,
siguiendo el gradiente osmótico creado por las sustancias absorbidas a la
sangre.
El paso del sodio al espacio intercelular
produce en éste una solución hipertónica que facilita el arrastre de agua como
consecuencia del gradiente osmótico creado. El agua pasa desde la
luz del intestino a través de las denominadas uniones estrecha, existentes
entre las membranas de células adyacentes (vía paracelular), o en menor
cuantía, a través del interior celular (vía transcelular). Las uniones
estrechas presentan una permeabilidad decreciente, desde el duodeno hasta el
colon, de tal modo que la participación de la vía paracelular en el transporte
de agua se va reduciendo a medida que se progresa a lo largo del
intestino. Una vez en el espacio intercelular, el agua, gracias al gradiente
de presión hidrostática creado, puede pasar a través de la membrana basal
(mucho más permeable que las uniones estrechas) hacia los capilares sanguíneos.
El cloro se absorbe en duodeno y yeyuno
por difusión pasiva siguiendo al Na+. En
el ileon y colon, realiza un cotransporte activo con el Na+, y
también un intercambio con el bicarbonato.
El potasio difunde a través de
canales y al interior celular por la bomba Na+/K+.
El calcio,se absorben diariamente de
200 a 300 mg mediante transporte activo transcelular, y transporte paracelular
pasivo.
El magnesio, se absorbe como el
calcio.
El fosfato, se absorbe alrededor del
65% mediante procesos pasivos, y también por un mecanismo acoplado al Na+,
utilizando un cotransportador.
El hierro, en su forma hemo, penetra en
las células intestinales mediante la formación de vesículas, seguido de
digestión enzimática y liberación del hierro libre que se combina
inmediatamente con la apoferritina para formar ferrolana. El hierro
en su forma no hemo, se libera de los alimentos y se solubiliza gracias a
la acidez del jugo gástrico entre otros, lo cual permite el paso de hierro férrico (Fe +++) a ferroso (Fe++).
6.6 Composición de las heces
A lo largo de un día se excretan de 100 a 150
gramos de materia fecal, de la que entre 30 y 50 gramos son sólidos y de 70 a
100 gramos son de agua. La materia sólida está formada por residuos de
materiales no digeribles (ej.: celulosa), pigmentos y sales biliares,
secreciones intestinales, principalmente mucus, células epiteliales descamadas,
bacterias (hasta 1/3 de los sólidos totales), y materiales inorgánicos: Ca++,
fosfatos, etc.
Las heces eliminadas diariamente a través del
recto contienen una cantidad de líquido inferior a los 200 ml. Cantidades
superiores determinan la aparición de diarrea, que puede tener diversos
orígenes. El fenómeno puede producirse, por ejemplo, por el uso de laxantes,
capaces de bloquear la actividad de la ATPasa Na/K y la absorción intestinal
de sodio. El cólera o determinadas infecciones bacterianas también pueden
originar diarrea al estimular la secreción de agua y electrolitos por las
criptas de Lieberkühn; por este mecanismo pueden llegar a perderse hasta 5‑10
litros de agua cada día, con grave riesgo para la vida.
6.6.1 Flora digestiva
El organismo humano contiene unos 100 billones de
bacterias (1014) que conviven de manera
simbiótica con él. Ha de observarse que esta población supone más de 10 veces
el número de células del cuerpo humano. Toda esta batería de microorganismos se
distribuye entre 400 especies diferentes, dando idea de la diversidad de este
ambiente ecológico.
La mayor parte se localiza en el colon donde se
miden concentraciones de 1012/ ml;
mientras que en el duodeno hay 104/ml.,
cantidad muy inferior, pero que indica la colonización bacteriana de todo el tubo
digestivo.
La denominada flora microbiana del aparato
digestivo desarrolla las siguientes funciones: 1. Metabólica. Ya que la hidrólisis de glúcidos
(fibra insoluble) y oligofructosacáridos, produce ácidos grasos de cadena corta
como:
Ácido butírico. Tiene un efecto trófico sobre
las células epiteliales del colon.
Ácido acético. Utilizado como nutriente por la
masa muscular.
Ácido propiónico. Utilizado como nutriente por el
hígado.
Además estas bacterias sintetizan vitaminas tales
como la biotina, ácido fólico, complejo B y vitaminas K y E. Añadidamente, el
pH que se genera en esta región facilita la absorción de Ca, Mg y Fe. Y una
función muy importante es la degradación del propio moco colónico, que en
ausencia de estas bacterias no es degradado correctamente y causa edemas en
esta región. 2. Efecto barrera frente a patógenos. Por un lado,
agotando los recursos nutritivos, de tal forma que no quedan para otras
especies; además controlando el pH del medio (acidificándolo) que imposibilita
la presencia de otros organismos de carácter patógeno. Y, por último, mediante
la liberación de bacteriocinas, que atacan otras bacterias. 3. Desarrollo, potenciación y modulación de la
respuesta inmune. El 80% de las células inmunocompetentes se encuentra en el
aparato digestivo, y en él se produce la mayor tasa de producción de
inmunoglobulinas (2-3 g de Ig A). En los primeros momentos de la vida, la
interacción de la flora microbiana con el sistema inmune produce la maduración
del mismo.
Hola, me llamo Jesús, nací en Badajoz el 3 de abril de 1996 y vivo en Valdebótoa que es un pueblo de Badajoz.
Tengo 16 años y no tengo hermanos.
Una de mis mayores aficiones son los animales.
uno de los que mas me gustan son los caballos,tengo cuatro caballos y monto a caballo aunque ya no concurso.
También me gustan los perros , la caza con perros y escopetas y el silvestrismo,pero menos que los caballos.
En el tema de los estudios quiero sacarme este año el graduado y luego empezar un módulo de agropecuaria ya que me gustan los animales.